CELULA. PERCY ZAPATA MENDO.
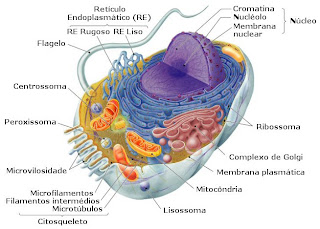
Célula 1 INTRODUCCIÓN Célula, unidad básica de la vida. La célula es la estructura más pequeña capaz de realizar por sí misma las tres funciones vitales: nutrición, relación y reproducción. Todos los organismos vivos están formados por células. Algunos organismos microscópicos, como las bacterias y los protozoos, son unicelulares, lo que significa que están formados por una sola célula. Las plantas, los animales y los hongos son organismos pluricelulares, es decir, están formados por numerosas células que actúan de forma coordinada. La célula representa un diseño extraordinario y eficaz con independencia de si es la única célula que forma una bacteria o si es una de los billones de células que componen el cuerpo humano. La célula lleva a cabo miles de reacciones bioquímicas cada minuto y origina células nuevas que perpetúan la vida. El tamaño de las células es muy variable. La célula más pequeña, un tipo de bacteria denominada micoplasma, mide menos de una micra de diá...